Back to Index
Page
An introduction to the palaeontology of rudist
bivalves
Rudist are a group of bivalves
which evolved during the Late Jurassic to Cretaceous and lived in warm, shallow
oceans of low latitudes. They became extinct at the Cretaceous/Tertiary
boundary. Most rudists have not much in common with 'normal' bivalves and
developed bizarre, occasionally large shells.
|
|
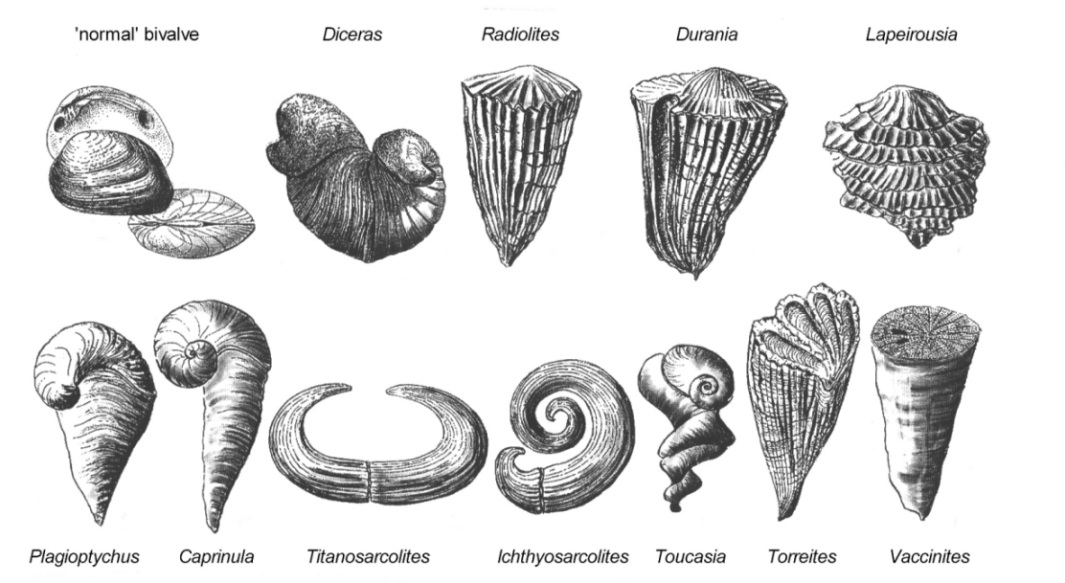
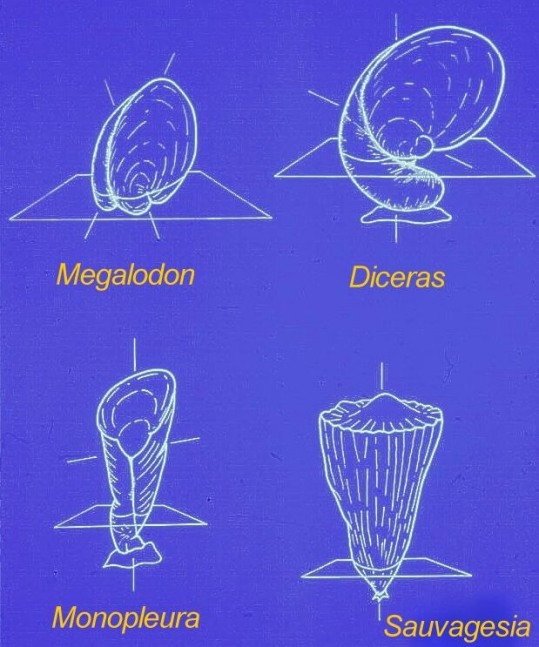
Different to other bivalves, one or
both valves are uncoiled which allowed for accretion of the shell along the
complete mantle margin and the construction of tubular shells.
|
|
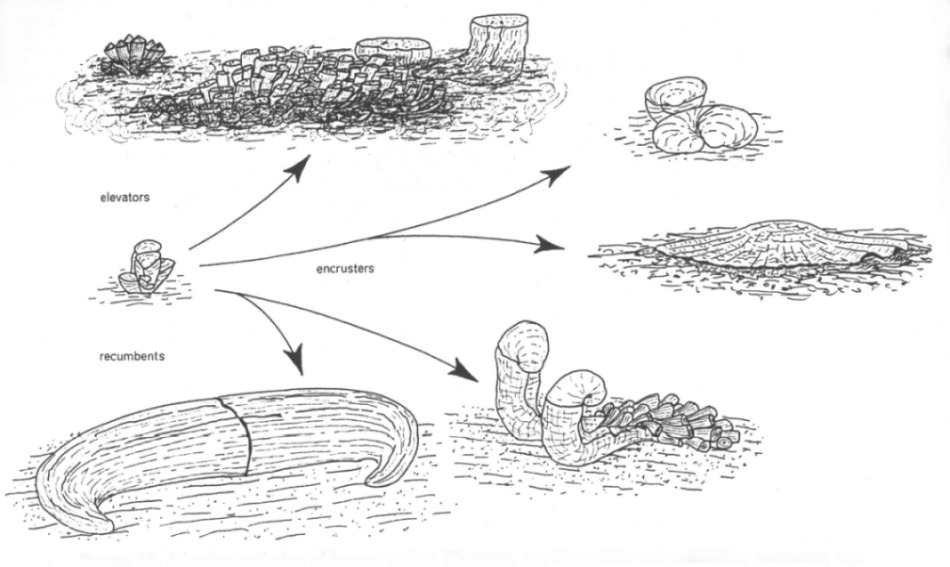
According to their life habit,
rudist morphotypes are classified as 'elevators', 'clingers' or 'recumbents',
each morphotype being adapted to specific environmental conditions (type of
sediment, sedimentation rate, current regime).
|
|
|

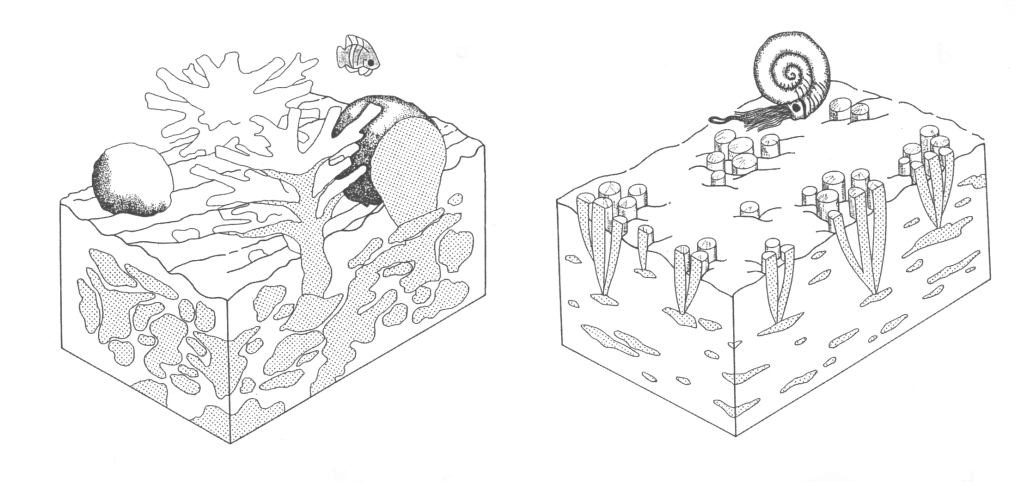
Although rudists are sometimes
considered as characteristic 'reef-builders' of the Cretaceous, several
important differences exist between typical coral-algal-hydrozoan reefs and rudist formations. Rudist formations are typically of low
relief and form more or less tabular bodies. Bound, wave-resistant fabrics were
uncommon, and elevator morphotypes were supported by sediment which accumulated
during their vertical shell growth. Consequently, growth fabrics of rudist
associations were constratal, in contrast to superstratal fabrics of modern
coral reefs.
|
|
Comprehensive information about rudist
formations are provided by, e.g., Gili et al. (1995), Höfling (1997), Kauffman & Sohl
(1973),
Philip (1984), Ross & Skelton
(1993), Sanders & Pons (1999).
The evolution
of rudist bivalves
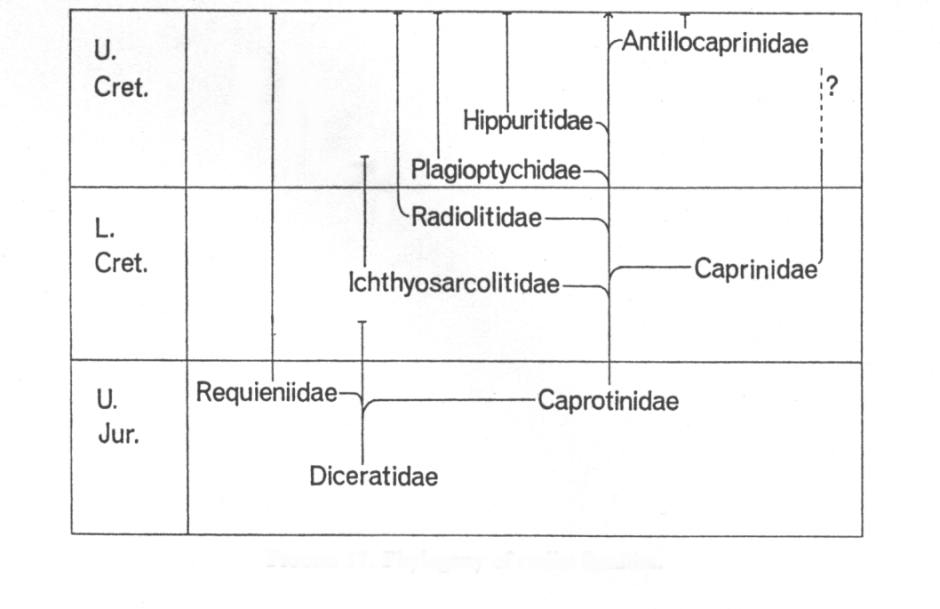
The Upper Jurassic - lower Cretaceous Diceratidae are
considered as the root stock of rudist bivalves. Uncoiling was only incipient,
and some genera were attached with the left valve while other were attached
with the right valve. The Requieniidae were particularly abundant in
Barremian-Lower Aptian Urgonian-type carbonate platforms. They were attached
with the left valve. The Caprotinidae and Polyconitidae, and all other,
derived families were attached with the right valve. The Caprinidae lived mainly as recumbents and have a
characteristic pattern of canals in one or both valves. Radiolitidae evolved since the Late
Aptian to become the most species-rich family among the group. Ichthyosarcolitidae
were particularly widespread during the Cenomanian, and have been rarely
recorded from older sediments. The Cenomanian/Turonian transition was critical
in the evolution of rudist. The Caprinidae and Ichthyosarcolitidae became
extinct, and the Hippuritidae
which dominated many Late Cretaceous rudist formations occurred for the first
time. Plagioptychidae
and Antillocaprinidae
are superficially similar to Caprinidae
but differ in the arrangement of myophores, supporting the adductor muscles.
The Antillocaprinidae are
restricted to the Americas and the Caribbean, and the Dictyoptychidae are only known
from the latest Cretaceous of the northeastern Afro-Arabian plate. The
extinction of rudist bivalves is hypothesised to have been stepwise during the
Maastrichtian, but the exact timing and pattern of extinction during the latest
Cretaceous is still enigmatic.
|
|
|
|
|
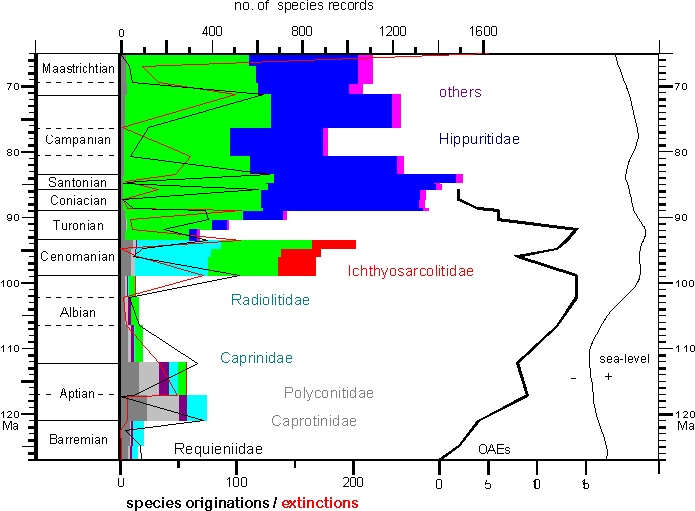
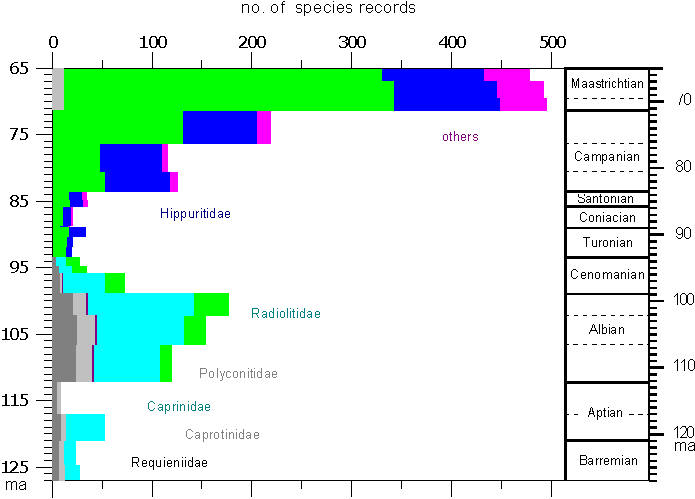
Most important for the distinction of rudist
families is the myocardinal arrangement and the structure of inner and outer
shell layers. Shells structures and ornaments are important for the subdivision
of families, genera and species.
|
|
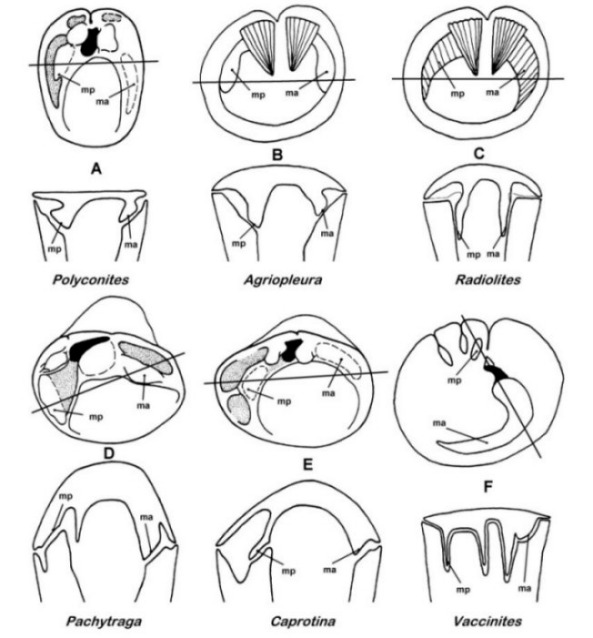
|
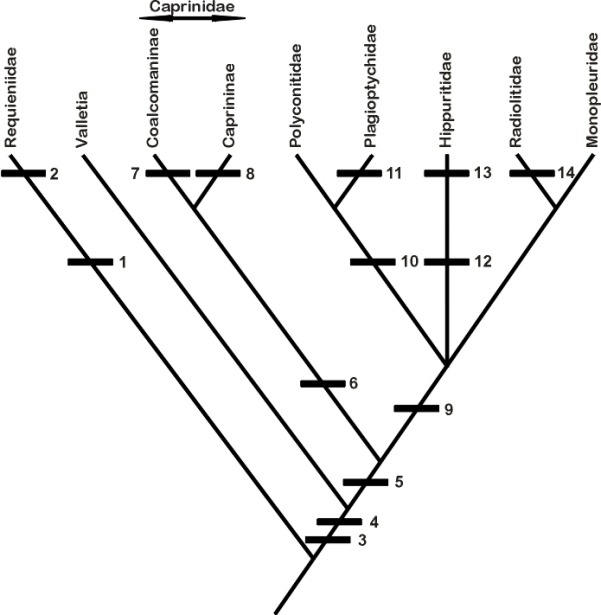
Tentative phylogeny of rudist bivales (Steuber 1999), a more detailed and rigorous cladistic
analysis is provided by Skelton & Smith (2000).
Derived characters: (1) left valve (LV) = attached valve; (2) thickened outer
shell layer; (3) right valve (RV) = attached valve; (4) two cardinal teeth in
LV, and one tooth, which is reduced in some taxa, in RV; (5) ligament
invaginated; (6) inclined to erect lamina in RV, running from central tooth
to posteroventral shell wall to demarcate omp’; (7) LV posterior myophore
(mp) insertion surface facing outwards onto mp´ of RV, the latter on
posterior shell wall; (8) LV mp insertion surface facing inwards onto outward
facing mp´ of RV, the latter on erect myocardinal lamina; (9) thick outer
shell layer in RV; (10) mp´ of RV on thickening of posterior shell (Polyconites), that may be raised to
extend parallel to the commissure or is even slightly tilted backwards (Horiopleura); (11) canals in inner
shell layer; (12) mp´ of RV tilted backwards, in tapering socket that receives
elongated mp of LV; (13) pores and canals in LV outer shell layer; (14)
cellular structure of outer shell layer in one or both valves. Phylogeny of
the Caprinidae d´Orbigny after Skelton and Masse (1998);
the Coalcomaninae Coogan include Retha
Cox, and the Caprininae d´Orbigny include Pachytraga
Paquier. |
{kind=link}
{kind=link}
Suggested reading on the evolution of rudists: Kauffman & Johnson (1988), Masse
& Philip (1986), Skelton (1978), Skelton & Smith (2000).
A wealth of information on all aspects of
rudist palaeontology can be found in the proceedings volumes of the
'International Conferences on Rudist Bivalves':
·
Geologica Romana, 28: 372
pp.; Roma (1992).
·
Alencáster, G. & Buitrón-Sánchez,
B.E.: (eds.), Number devoted to the Third international
Conference on Rudists. -- Revista mexicana de Ciencias geológicas, 12/2 (for 1995):
316 pp.; Mexico City (1996).
·
Masse, J.-P. & Skelton,
P.W.: (eds.), Quatrième Congrès international sur les Rudistes.
-- Geobios, Mémoire
spéciale, 22: 427 pp.; Lyon (1998).
·
A Bibliography is also available:
·
Steuber,
T. & Löser, H. (1996):
Jurassic-Cretaceous rudists (Mollusca, Hippuritacea) - Bibliography and data
base 1758-1994.- Neue
Paläontologische Abhandlungen, 1:
123 pp., Diskette; Dresden.
- Steuber, T. &
Löser, H. (1998):
Jurassic-Cretaceous rudists (Mollusca, Hippuritacea) - Bibliography
1758-1994, 1st supplement.- Coral Research Bulletin, 6: 147-182, Diskette; Dresden.